WCRP 111 : Proceedings of the third PMIP Workshop
PALAEOENVIRONMENTAL DATA SETS AND MODEL EVALUATION IN PMIP
Sandy P. Harrison
Max Planck Institute for Biogeochemistry,
P.O. Box 100164, D-07701 Jena, Germany
Sandy.Harrison@bgc-jena.mpg.de
1. INTRODUCTION
Physically-based models provide the unique means to predict the likely
impacts of anthropogenic changes in atmospheric composition and land
use (Intergovernmental Panel on Climate Change, 1997). The accurate
simulation of current climate is an important benchmark but does not
guarantee that a model will simulate climate changes correctly
(Joussaume et al., 1999). It difficult to evaluate model performance
solely on the basis of the instrumental record because the changes in
climate since the middle of the last century have been relatively
modest (e.g. Mann et al., 1995; Tett et al., 1999). Evaluating model
performance under the extreme climate conditions of the last glacial
maximum (LGM: ca 21,000 calendar yr B.P., equivalent to ca 18,000 14C
yr B.P.) and the mid-Holocene (ca 6000 yr B.P.) provides an
opportunity to evaluate how models respond to larger changes in
forcing, and ultimately provides a key credibility test for modelling
the future (Joussaume and Taylor, this volume). However, such an
evaluation is crucially dependent on the existence of
spatially-explicit data sets which can be compared with output from
the model simulations. Thus, one goal of the Palaeoclimate Modelling
Intercomparison Project (PMIP) has been to foster the creation of
well-documented, spatially-explicit data sets explicitly designed for
use in model evaluation. Although the construction of
palaeoenvironmental data sets for model evaluation began prior to PMIP
(see e.g. Street and Grove, 1976; Peterson et al., 1979; COHMAP
Members, 1988; Wright et al., 1993), PMIP has played a key role in
stimulating the continued development and improvement of such data
sets and has been instrumental in the creation of two new data sets:
the BIOME 6000 data set (Prentice and Webb, 1998; Prentice et al., in
press) and the 21ka Tropical Terrestrial Data Synthesis (Farrera et
al., 1999).
2. PMIP APPROACHES TO DATA-MODEL COMPARISONS
Two basic approaches to comparing observations and simulations have
been used within PMIP: inverse techniques and forward-modelling
techniques. Inverse and forward-modelling techniques provide
complementary approaches to data-model comparisons. Inverse methods
are particularly useful when the geological data are abundant. The
forward modelling approach maximizes the use of relatively sparse data
sets for model evaluation.
In the inverse technique, palaeodata are translated into climatic
parameters via statistical techniques (e.g. transfer functions,
response surfaces or modern analog analyses: see Guiot, 1991 for a
review of these techniques) which rely on establishing empirical
relationships between modern observations and environments. Initially,
inverse techniques were used to reconstruct standard climate
parameters (e.g. mean July temperature, mean January temperature, mean
annual precipitation). This approach yields robust reconstructions,
with acceptably small error bars, in circumstances where the sensor is
directly controlled by aspects of the climate that are well correlated
with the standard climate parameters. This is not always the case. For
example, one of the most important controls on vegetation growth is
the accumulated temperature sum during the growing season (growing
degree-days, GDD: Prentice et al., 1992). GDD is well correlated with
mean July temperature when July temperatures are <18°C but not in
warmer climates, and nor is GDD exactly predictable as an empirical
combination of July and January temperatures (Kohfeld and Harrison,
2000). Furthermore, when GDD is used to predict vegetation limits it
is being used as a proxy for net primary production (NPP) which can be
influenced by low atmospheric CO2 concentrations (Jolly and Haxeltine,
1997; Street-Perrott et al., 1997; Cowling, 1999; Cowling and Sykes,
1999). In order to avoid such oversimplifications of the
climate-vegetation relationship, PMIP has strongly encouraged
reconstructions of non- standard climatic variables that are more
closely related to the underlying controls on specific
palaeoenvironmental indicators. Evaluation data sets derived from
inverse reconstructions of climatic or bioclimatic parameters for the
PMIP 6000 yr B.P. and LGM timeslices have been constructed for a
number of continental-scale regions from which there is abundant
fossil data, including Europe (Guiot et al., 1993; Cheddadi et al.,
1997; Peyron et al., 1998), eastern North America (Webb et al., 1993;
Williams et al., this volume), Russia (Tarasov et al., 1999; Tarasov
et al, in press), China (Yu and Qin, 1997) and the tropics (Farrera et
al., 1999). Guiot et al. (submitted; see also Guiot et al., this
volume) have used explicit inversion of a forward model to reconstruct
palaeotemperatures from LGM pollen data taking into account the
physiological effects of low atmospheric CO2 concentrations at the
LGM.
In the forward-modelling approach, process-based models are used to
predict the response of palaeoenvironmental indicators
(e.g. vegetation, hydrology) to the simulated climate. The predicted
response is then compared directly with palaeo-observations. Although
additional uncertainties about the cause of mismatches between
simulations and observations can be caused by the use of a second
model, forward modelling has been used within the PMIP project to
facilitate comparisons with terrestrial vegetation data and with lake
data. Thus, we have used terrestrial biosphere models from the BIOME
model family (e.g. BIOME1: Prentice et al., 1992; BIOME3: Haxeltine
and Prentice, 1996; BIOME4: Kaplan et al., in prep.) to simulate the
distribution of major vegetation types (biomes) (see e.g. Harrison et
al., 1995; Kutzbach et al., 1996; TEMPO, 1996; Harrison et al., 1998;
Kutzbach et al., 1998; Harrison et al., in prep.) for direct
comparison with biomes reconstructed from palaeovegetation data from
the BIOME 6000 data set (see below). Terrestrial hydrological models,
which predict the surface area of lakes and wetlands, and river
discharge, using land-surface morphology and simulated runoff,
precipitation, and evaporation (e.g. HYDRA: Coe, 1998; Coe, in press)
have been used in a similar manner and directly compared with
palaeolake area and river discharge data (Coe and Harrison, this
volume) derived from the Global Lake Status Data Base (see below).
3. GLOBAL PALAEOENVIRONMENTAL EVALUATION DATA SETS
There are three types of data that have been widely used for model
evaluation within PMIP: the Global Lake Status Data Base, the BIOME
6000 datasets, and the 21ka Tropical Terrestrial Data Synthesis.
3.1 The Global Lake Status Data Base
Lakes respond in a simple and well-understood fashion (see Cheddadi et
al., 1997) to changes in the balance of precipitation and evaporation
(P-E) over the lake and catchment by changing in volume (and hence
normally monotonically in area or level). In overflowing lakes in
humid regions, runoff from the catchment primarily determines the
equilibrium lake level and hence the magnitude of lake discharge.
Sustained changes in lake discharge are only produced when lake volume
is also changed. In closed lakes from semi-arid regions, the area of
the lake simply represents an equilibrium between runoff from the
catchment and the water deficit over the lake. The Global Lake Status
Data Base (GLSDB: Qin et al., 1998; Kohfeld and Harrison, 2000) is a
long-standing international effort to compile the geomorphic and
biostratigraphic data (see Harrison and Digerfeldt, 1993 for
discussion of types of evidence) for changes in lake level, area, or
volume (collectively referred to as lake status), in order to document
changes in regional water balance during the last 30,000
years. Developed with data-model comparisons as a primary objective,
the GLSDB builds on the earlier Oxford Lake Level Data Base (Street
and Grove, 1976; Street-Perrott and Harrison, 1985; Street-Perrott et
al., 1989), and contains data both from closed-basin lakes in now-arid
regions and from currently overflowing lakes in temperate and wet
tropical regions (Yu and Harrison, 1996; Tarasov et al., 1994, 1996;
Jolly et al., 1998a; Qin et al., 1998; Kohfeld and Harrison, 2000; see
also Yu et al., this volume).
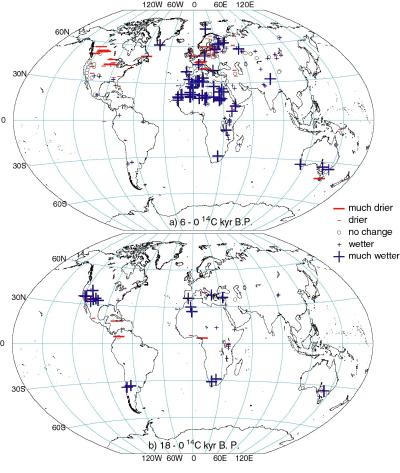
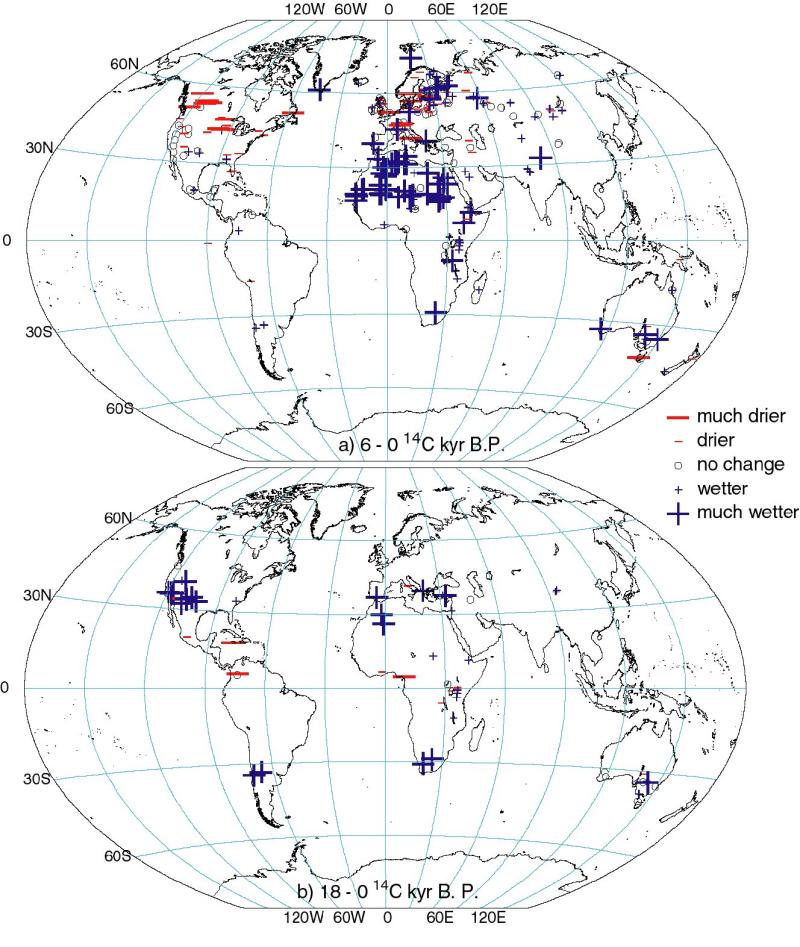
Lake status data from the GLSDB for 6000 yr B.P. (Figure 1a) show that
conditions were wetter than today across northern Africa, the Arabian
Peninsula, northern India, and southwest China, indicating expansion
of the Afro-Asian summer monsoons. Conditions were slightly wetter or
comparable to today in central America, SW USA, central Eurasia, in
the mid-latitudes of the southern hemisphere, and in the high northern
latitudes. Exceptions to this pattern, where lakes show conditions
were drier than today, are seen in interior North America and western
Europe.
Lake status data from the GLSDB for the LGM (Figure 1b) show
conditions as dry or drier than today over much of the world as a
result of the generally weaker than present hydrological cycle.
However, lakes register wetter conditions in western North America and
the circum- Mediterranean region, and in the Southern Hemisphere
mid-latitudes.
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
Figure 1 - Changes in lake status at (a) 6000 14C yr
B.P. and (b) at 18,000 14C yr B.P. (~21,000 calendar yr
B.P.) compared to present. Only sites with dating control of <= 4
for 6000 14C yr B. P., and < 7 for 18,000 14C
yr B.P. are included.
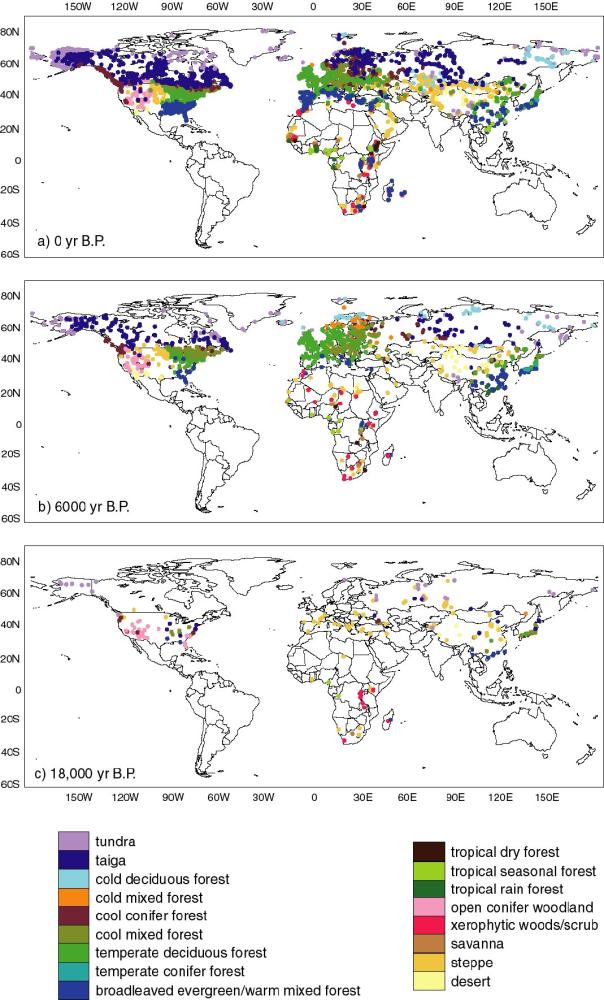
3.2. BIOME 6000
The Palaeovegetation Mapping Project (known as BIOME 6000: Prentice
and Webb, 1998) was established in 1994 with the aim of developing
global palaeovegetation data sets for the LGM (here defined as 18,000
± 2000 14C yr B.P.) and the mid-Holocene (6000 ± 500 14C
yr B.P.) which could be explicitly used to evaluate palaeoclimate
model simulations. Broadscale vegetation types (biomes) are
reconstructed from pollen or plant-macrofossil data using a
standardized, objective method (biomization) based on plant functional
types (PFTs: Steffen et al., 1992; Prentice et al., 1996). Plant taxa
are first assigned to PFTs, and then the set of PFTs that can occur in
each biome is specified. The allocation of pollen or
plant-macrofossil assemblages to biomes is made on the basis of an
affinity-score procedure which takes into account both the diversity
and the abundance of taxa belonging to each PFT in the sample.
Extensive tests using modern surface samples have shown that the
method is capable of reproducing natural vegetation patterns even in
regions heavily impacted by human activities (Figure 2a).
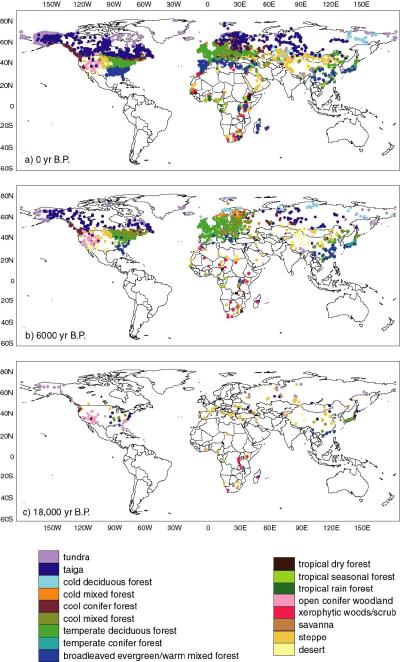
The BIOME 6000 data set for 6000 yr B.P. (Figure 2b) shows that the
Arctic forest limit was north of its present position in the Mackenzie
Delta region (Edwards et al., in press), Europe (Prentice et al.,
1996) and western and central Siberia (Texier et al., 1997; Tarasov et
al., 1998), unchanged in Beringia (Edwards et al., in press), northern
Canada and Keewatin (Williams et al., in press) and south of its
present position in Quebec-Labrador (Williams et al., in press). The
northward expansion of northern temperate forest zones was more
dramatic than the relatively modest change in the Arctic forest limit.
Warmer winters (as well as summers) are required to explain some of
these shifts in northern temperate forests (Prentice et al., in
press). Temperate deciduous forests were greatly extended in Europe,
southwards into the Mediterranean region as well as to the north
(Prentice et al., 1996). Steppe vegetation occurred in areas occupied
today by forests in North America in response to drier conditions
(Williams et al., in press), but forest biomes encroached on the
present-day steppe in southeastern Europe and Central Asia (Tarasov et
al., 1998). Enhanced monsoons extended forest biomes inland in China
(Yu et al., 1998; Yu et al., in press; see also Yu et al., this
volume) and Sahelian vegetation into the Sahara, while the African
rainforest was reduced (Jolly et al., 1998a, b) consistent with a more
seasonal climate in the equatorial zone.
The BIOME 6000 data set for the LGM (Figure 2c) shows that cold and
dry conditions at the LGM favoured extensive tundra and steppe
vegetation (Prentice et al., in press). Northern hemisphere boreal
and temperate forest biomes were displaced southward and fragmented.
Tropical moist forests in Africa were also reduced in extent (Elenga
et al., in press). Open conifer woodlands were more extensive than
today in southwestern North America, however, indicating conditions
wetter than today in this region (Thompson and Anderson, in press) in
contrast to the general trend towards aridity.
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
Figure 2 - Biome reconstructions for (a) modern, (b) 6000
14C yr B.P. and (c) 18,000 14C yr B.P. (from
Prentice et al., in press).
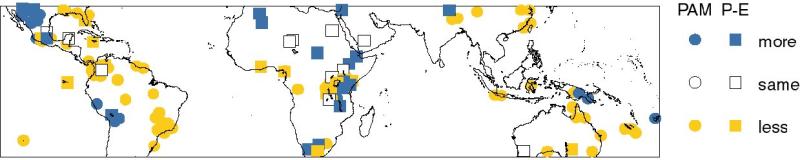
3.3. 21 ka Tropical Terrestrial Data Synthesis
The 21 ka Tropical Terrestrial Data Synthesis (Farrera et al., 1999)
contains quantitative reconstructions of mean temperature of the
coldest month (MTCO) and mean annual ground temperature (MAT), and
qualitative reconstructions of plant-available moisture (PAM) and
runoff (P-E) from radiocarbon-dated terrestrial sites between 32°N
and 33°S with records for 18,000±2000 14C yr B.P. The
dataset combines multiple indicators of quantitative changes in land
surface temperature (pollen and plant macrofossil records of MTCO, and
noble gas and speleothem records of MAT) and qualitative indicators of
moisture balance parameters (pollen and plant macrofossil records of
PAM and lake status records of P-E). The use of multiple indicators
allows the consistency of the temperature and moisture balance
reconstructions to be evaluated.
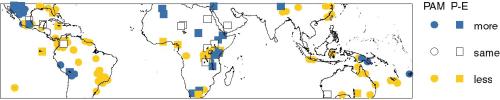
PAM and P-E are complementary measures of the water balance: runoff is
the water that is not used by plants (Prentice et al., 1993).
However, in regions where there are both kinds of data, the observed
changes at the LGM within the 32°N and 33°S band are in the same
direction (Figure 3). Both indicators show conditions drier than
today at the LGM across most of the tropics and subtropics, except in
the Great Basin, E. Africa and at high elevations in South America and
Papua New Guinea. Lakes in northern Africa, supplied by runoff from
the Atlas Mountains (Street-Perrott et al., 1989), also show wetter
conditions.
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
Figure 3 - Reconstructions of PAM (circles) and P-E (squares) at the
LGM, from Farrera et al. (1999).
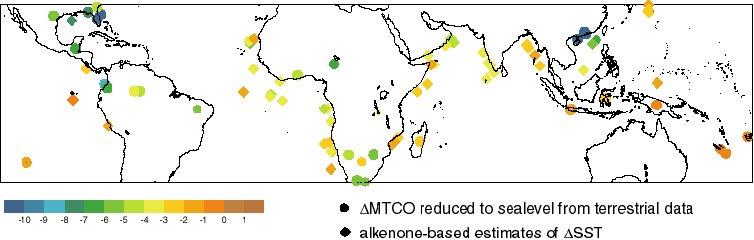
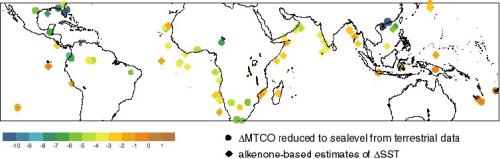
The LGM changes in MAT and MTCO are broadly similar at low latitudes
because the low seasonality of insolation in the tropics offers little
scope for seasonal changes in temperature, but the two variables
differ systematically in the northern subtropics as seasonal
differences become greater. Both variables indicate that temperatures
in the tropics were on average 2.5-3°C cooler than today at modern sea
level (Figure 4). However, the magnitude of the cooling is not
spatially uniform even in the lowlands: the Neotropical region
(Central and northern South America) cooled by 5-6°C; the cooling at
circum-Indian Ocean sites (South and East Africa, India and Indonesia)
was ca 2-3°C; the cooling was less than 2°C in the circum-Pacific
region (Papua New Guinea, and the islands of the western Pacific
Ocean). Spatial differences are also apparent in the lapse rates at
tropical sites. Lapse rates were steeper by ca 2?/km in the
circum-Pacific region, by ca 1?/km in the Indian Ocean region, while
there was no discernable change in lapse rate in the Neotropical
region.
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
Figure 4 - Comparison of DELTA(MTCO) reduced to sea level (circles)
based on lowland terrestrial from the Farrera et al. (1999) data set
and alkenone-based estimates of DELTA(SST) (diamonds) from the TEMPUS
compilation.
4. EVALUATION OF THE PMIP SIMULATIONS USING PALEOENVIRONMENTAL
DATA SETS
These three global data sets, and the various regional climate
reconstructions described above, have been widely used to evaluate the
PMIP baseline and complementary experiments (see e.g. Yu and Harrison,
1996; Texier et al., 1997; Broström et al., 1998; Vettoretti and
Peltier, 1998; Joussaume et al., 1999; Masson et al., 1999; Pinot et
al., 1999; Guiot et al., in press; Bonfils et al., this volume;
Braconnot et al., this volume; Coe and Harrison, this volume; Guiot et
al., this volume; Kageyama et al., this volume; Williams et al., this
volume; Yu et al., this volume). The aim of this section is not to
document all that we have learnt from such evaluations but rather to
provide an example of how the data can be used as a benchmark for
model evaluation. Specifically, we will show how data from northern
Africa has been used as a benchmark for the evaluation of PMIP
baseline and complementary simulations of the mid- Holocene monsoon
system, and then how the new data from the tropics has been used to
evaluate PMIP LGM simulations.
4.1 Evaluation of the PMIP 6000 yr B.P. experiment
The expansion of the area influenced by the Afro-Asian summer monsoons
at 6000 yr B.P. is one of the most striking features shown by
palaeoenvironmental data. The mechanisms underlying the enhancement of
the African monsoon have been known for some time (Kutzbach and
Street-Perrott, 1985; COHMAP Members, 1988). The PMIP simulations
(Joussaume et al., 1999) confirm that the orbitally-induced
enhancement of northern-hemisphere summer insolation at 6000 yr
B.P. resulted in increased heating over the northern hemisphere
continents and thus intensified the thermal contrast between the land
and the ocean. The increased heating over northern Africa resulted in
the northward displacement of the ITCZ and hence of the monsoon front
over northern Africa, while the enhanced land-sea contrast increased
the flux of moisture from the ocean to the continent. However,
comparisons of the spatial patterns in the simulated P- E fields with
lake data from the GLSDB (Yu and Harrison, 1996) indicate that the
PMIP simulations consistently underestimate the northward shift in the
monsoon front. Similarly, BIOME3 simulations made with output from the
PMIP simulations consistently fail to reproduce the observed northward
shift in the Sahara/Sahel boundary (Harrison et al., 1998). The
precipitation required to generate the observed latitudinal
distribution of steppe (grassland) vegetation in northern Africa at
6000 yr B.P. has been estimated using a combination of
forward-modelling and inverse techniques. Joussaume et al (1999)
showed that the PMIP simulations underestimate the required
precipitation at ca 23°N by at least 100mm (Figure 5a), i.e. by ca
50% of the minimum amount required to support steppe. When output from
the PMIP experiments is used to simulate the extent of lakes across
northern Africa using the HYDRA model, the observed area of Lake Chad
(350,000 km2) is significantly underestimated by all of the
models (Coe and Harrison, this volume). Thus, data-model comparisons
show that the PMIP simulations consistently underestimate both (1) the
northward shift in the monsoon belt shown by palaeoenvironmental data,
and (2) the magnitude of the precipitation required to produce the
observed lake and vegetation changes in northern Africa.
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
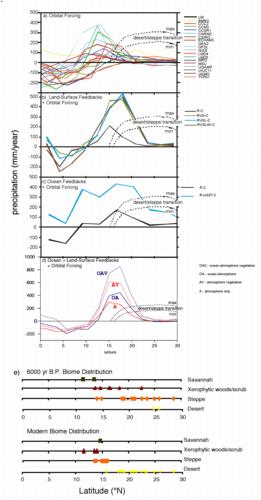
Figure 5. Zonally-averaged simulated annual precipitation anomalies
(6000 cal. yr B.P. - control) vs latitude for northern Africa (land
grid cells, 20°W-30°E). Precipitation anomalies include the
effects of: (a) radiative forcing (R) alone for the 18 climate models
participating in PMIP (Joussaume et al., 1999); (b) radiative forcing
plus land-surface feedbacks (soil: S, vegetation: V, lakes: L, and
wetlands: W) simulated using CCM3 (Broström et al., 1998); (c)
radiative forcing plus ocean feedbacks (DELTA(SST)) for an
asynchronous coupling of GENESIS2 and MOM1 (Kutzbach and Liu, 1997).
(d) radiative forcing plus ocean- and vegetation-feedbacks from an
asynchronous coupling of the IPSL AOGCM and BIOME1 (Braconnot et al.,
1999). The hatched lines in (a-d) represent upper and lower estimates
of the additional precipitation (excess over modern) required to
support the steppe vegetation observed in northern Africa at 6000
14C yr (see Joussaume et al., 1999). (e) Latitudinal
distribution of biome types (desert, steppe, xerophytic woods/scrub,
and savannah) for 6000 14C yr and modern over the
longitudes 20W-30E (Jolly et al., 1998b; Joussaume et al., 1999).
4.2. Evaluation of Complementary 6000 yr B.P. PMIP Experiments
In the baseline PMIP simulations land-surface conditions and ocean
surface temperatures were prescribed to be the same as today
(Joussaume and Taylor, this volume). PMIP has also investigated the
importance of feedbacks associated with changes in land-surface or
ocean- surface conditions e.g. for the simulation of the African
monsoon, using palaeodata as a benchmark. A sensitivity study which
sequentially examined the response to realistic changes in (a)
vegetation and soils, (b) the extent of lakes and (c) the extent of
wetlands across northern Africa showed that land-surface feedbacks
amplify the orbitally-induced monsoon by increasing precipitation
during the peak of the monsoon season and extending the monsoon season
by 2-3 months, resulting in a shift of the monsoon front ca 300 km
further north than by insolation changes alone (Broström et al.,
1998). Nevertheless, comparisons with the benchmark data (Figure 5b)
show that the feedbacks were insufficient to produce the full observed
northward expansion of grasslands into regions occupied today by
desert. Similar evaluations (not shown) of experiments in which the
land-surface feedbacks are simulated either through asynchronous-
coupling of an equilibrium vegetation model with an AGCM (e.g. Texier
et al., 1997; Claussen and Gaylor, 1997; de Noblet et al., in press)
or using dynamically coupled atmosphere-vegetation models
(e.g. Doherty et al., in press) confirm that land-surface feedbacks
are insufficient to explain the full observed expansion of the African
monsoon.
Sensitivity experiments is which sea-surface temperatures in the
tropical and subtropical Atlantic were prescribed to be 1-3°C
colder than today show ocean feedbacks increase monsoon rainfall in
the Sahara/Sahel by enhancing evaporation from the ocean surface and
increasing moisture flux to the continent (Texier et al., 2000). In
asynchronously coupled atmosphere-ocean experiments (Kutzbach and Liu,
1997), maximum monsoon precipitation shifts northward to 15- 20°N
and precipitation over northern Africa increases by 25% compared to
simulations made with prescribed modern SSTs. However, the
precipitation increase induced by the combined effect of orbital
forcing and ocean feedbacks is still not enough to support the
observed grassland vegetation north of 23°N (Figure
5c). Simulations using fully-coupled atmosphere-ocean models
(e.g. Hewitt and Mitchell, 1998; Otto-Bleisner, 1999; Braconnot et
al., in press; Braconnot et al., this volume) confirm that
ocean-surface feedbacks produce a significant enhancement of the
orbitally-induced changes in the African monsoon but again the changes
are apparently insufficient to explain its observed expansion.
PMIP participating groups have begun to explore whether synergistic
feedbacks involving land-atmosphere-ocean interactions are involved
the observed expansion of the African monsoon during the mid-Holocene
using asynchronously or fully-coupled ocean-atmosphere-vegetation
models (Ganopolski et al., 1998; Braconnot et al., 1999) and benchmark
palaeoclimate data sets (Figure 5d).
4.3 Evaluation of the PMIP 21,000 yr B.P. Experiments
Palaeoenvironmental data show that conditions were colder and, except
in regions influenced by the southward displacement of the Westerlies
by the Laurentide Ice Sheet, considerably drier than today (COHMAP
Members, 1988; Kohfeld and Harrison, 2000). In the tropics, land-based
estimates indicate temperatures were on average 2.5-3°C cooler
than today at modern sea level, although there were large meridional
differences in the magnitude of the cooling ranging from 1- 2°C in
the circum-Pacific to 5-6°C in the Neotropics (Farrera et al.,
1999). These land-based estimates of cooling at sea level are
consistent with reconstructions of the cooling based on marine
alkenone (Rostek et al., 1993; Schneider et al., 1995; Bard et al.,
1997; Rosell-Melé et al., 1998; Sonzogni et al., 1998) data
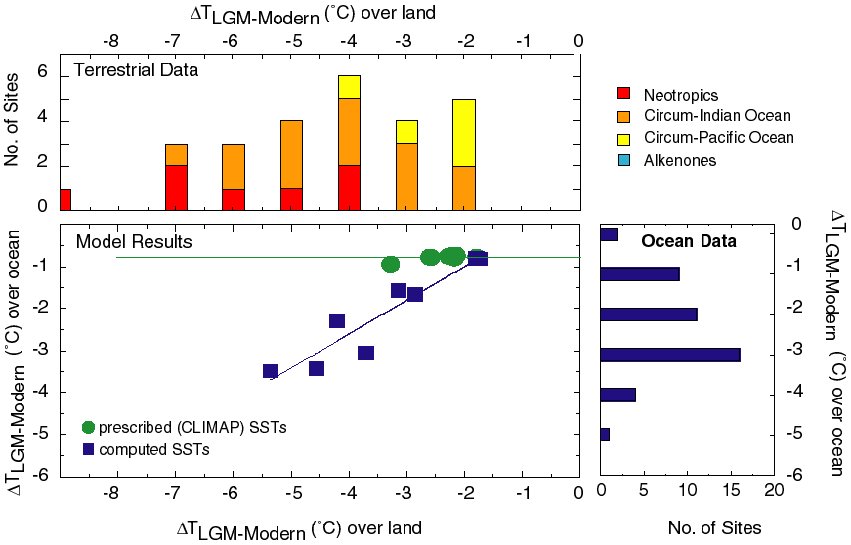
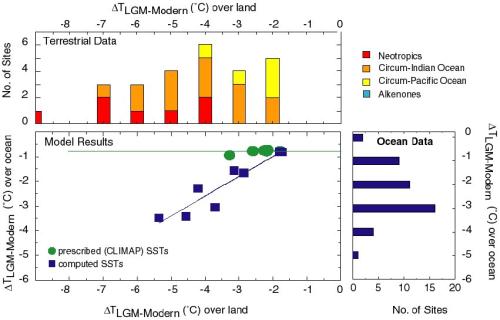
(Figure 4). The PMIP simulations with prescribed CLIMAP SSTs show only
a weak cooling in the tropics (Figure 6) and an increase in
continental aridity that is less than shown by the
observations. Analyses of the tropical response of the PMIP models
(Pinot et al., 1999) thus confirm Rind and Peteet's (1985) conclusions
that AGCMs with prescribed CLIMAP SSTs underestimate the observed
magnitude of tropical cooling at the LGM inferred from palaeodata. The
simulations with computed SSTs show a wide range in the magnitude of
tropical cooling, with some models underestimating and others
overestimating the mean cooling (Pinot et al., 1999). However, at
least some of the simulations made with computed SSTs produce a
tropical cooling that is consistent with the temperature changes shown
by terrestrial (Figure 4; Farrera et al., 1999) and marine alkenone
(Figure 4; Rostek et al., 1993; Schneider et al., 1995; Bard et al.,
1997; Rosell-Melé et al., 1998; Sonzogni et al., 1998) data.
Most of the models produce a rather uniform zonal cooling, which is
inconsistent with the distinct differences in the magnitude of the
cooling shown by data from e.g. the Neotropics and the western Pacific
(Figure 6). However, the UKMO model produces a meridional patterning
in tropical temperature changes consistent with the terrestrial and
the alkenone data (Figure 6).
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
Figure 6. Comparison of the simulated tropical (30°N-30°S)
cooling over land as a function of the intensity of the tropical
cooling over the ocean as simulated by the PMIP models with prescribed
(green circles) and computed (blue squares) SSTs (from Pinot et al.,
1999), and reconstructions of the land cooling based on terrestrial
data (from Farrera et al., 1999) and ocean cooling based on alkenone
data (from the TEMPUS data set).
4.4. Evaluation of Complementary LGM PMIP Experiments
Land-surface conditions in the PMIP LGM simulations were prescribed to
be the same as in the modern control simulation. Thus these
experiments (like those for 6000 yr B.P.) neglect the potential role
of land-surface feedbacks on glacial climates. Levis et al. (in press)
have shown that vegetation feedbacks, and specifically a simulated
reduction in tropical forests, in a coupled
atmosphere-vegetation-mixed layer ocean model LGM simulation cooled
the tropics by several degrees. Precipitation in the wet tropics and
in eastern Asia was substantially reduced in this simulation, in good
agreement with the widespread reduction of PAM in the tropics (Farrera
et al., 1999) and the replacement of forests with desert and steppe
vegetation in China shown in the BIOME 6000 data set (Yu et al., in
press).
Vegetation distribution and structure play an important role in
determining dust emissions, and the known changes in vegetation cover
at the LGM could have caused changes in atmospheric dust loading that
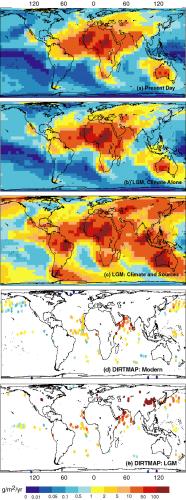
might have had a significant impact on the LGM climate. Ice cores,
marine sediments and loess deposits show that glacial periods were
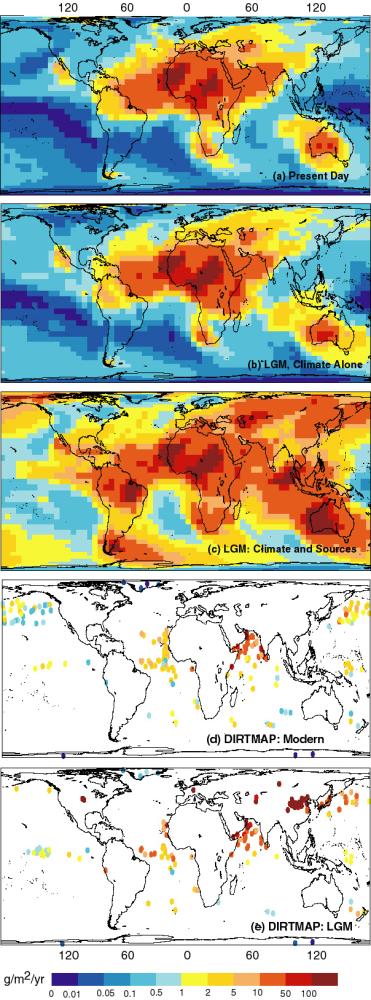
2-20 times dustier than interglacial periods (Figure 7a: Kohfeld and
Harrison, 2000; Harrison et al., in press). Mahowald et al. (1999)
have examined the relative importance of the direct effects of the LGM
climate (increased wind strengths, decreased hydrological cycle) and
the indirect effects of the changed climate through increasing dust
source areas on the production and distribution of dust using climate
output from one of the PMIP simulations (ECHAM 3.2, driven by CLIMAP
SSTs: Lorenz et al., 1996). They estimated (1) potential dust source
areas using a terrestrial biosphere model (BIOME3: Haxeltine and
Prentice, 1996); (2) entrainment using a simple source function
related to soil texture and wind velocity (Marticorena and Bergametti,
1996; Schulz et al., 1998); and (3) transport using an atmospheric
transport model (TM3: Heimann, 1995). Mahowald et al. (1999) showed
that, although changes in wind strength and precipitation could
explain the increased dust deposition registered in marine cores off
western Africa (7b), the expansion of source area in high-northern
latitudes, Asia and Patagonia are required to account for the 2-5 fold
increase in the global atmospheric dust content and 20-fold increase
at the poles at the LGM. (Figure 7c). An offline assessment of the
radiative impact of the relatively realistic dust fields simulated by
Mahowald et al. (1999) show that the simulated dust loading caused a
reduction in forcing (-0.4 to -1.1 W m-2) in the tropics
(Harrison et al., in press; Claquin et al., submitted). This forcing
represents an effect of comparable magnitude to the tropical cooling
effect of low atmospheric CO2 concentrations (Hewitt and
Mitchell, 1997).
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)
Figure 7 - Dust deposition simulated (a) under modern climate
conditions, (b) in response to LGM climate changes, and (c) in
response to LGM climate and expanded source areas (from Mahowald et
al., 1999) compared with (d) modern dust deposition and (e) LGM dust
deposition, based on geological records from the DIRTMAP data base
(Kohfeld and Harrison, 2000; Harrison et al., in press).
IMPLICATIONS FOR THE FUTURE
The PMIP community, through systematic comparisons of model
simulations against benchmarks provided by regional or global
syntheses of palaeoenvironmental data, has demonstrated that the
observed large changes in mid-Holocene and LGM climates cannot be
simulated without explicitly considering ocean- and land-surface
feedbacks. There may be a number of other important potential
feedbacks (e.g. radiative forcing by mineral aerosols). COHMAP
results (Kutzbach et al., 1998) indicating that the apparent
mismatches between observed and simulated climate changes during the
transition from glacial to interglacial conditions are greater than at
either the LGM or the mid-Holocene suggest that the incorporation of
these feedbacks may be even more important in attempts to simulate
times of rapid climate change when there is a strong disequilibrium
between insolation and other conditions. The fully- coupled
ocean-atmosphere-vegetation models (OAVGCMs) that will be required to
explore these issues will need to be rigorously benchmarked against
palaeoenvironmental data. The PMIP project therefore remains committed
to improving existing data sets, developing better analytical tools
and encouraging additional syntheses of palaeoenvironmental data in
order to be able to evaluate the models that will subsequently be used
to simulate potential future climate changes.
ACKNOWLEDGEMENTS
The GLSDB is sponsored by IGBP through its programme element Past
Global Changes (PAGES) Palaeo-Mapping Project (PMAP). BIOME 6000 is
sponsored by IGBP through its programme elements Global Analysis,
Intercomparison and Modelling (GAIM), the Data and Information System
(DIS), Global Change and Terrestrial Ecosystems (GCTE) and PAGES. The
members of the BIOME 6000 project are: Afanas'eva, N.B., Ager, T.,
Anderson, K., Anderson, P.M., Andrieu, V., Andreev, A.A., Ballouche,
A., Bartlein, P.J., de Beaulieu, J.L., Bengo, M., Berezina, N.A.,
Bezusko, L.G., Bezusko, T.V., Bigelow, N.H., Blyakharchuk, T.A.,
Bolikhovskaya, N.S., Bonnefille, R., Bottema, S., Brénac, P.,
Brubaker, L.B., Buchet, G., Burney, D., Bykova, G.V., Cheddadi, R.,
Chen, X., Chernavskaya, M.M., Chernova, G.M., Cwynar, L.C., Dorofeyuk,
N.I., Dirksen, V.G., Edorh, T., Edwards, M.E., Eisner, W.J., Elenga,
H., Elina, G.A., Elmoutaki, S., Filimonova, L.V., Glebov, F.Z., Guiot,
J., Gunova, V.S., Hamilton, A.C., Han, H., Harrison, S.P., Hu, F.-S.,
Huang, C., Huntley, B., Jolly, D., Jonson, H., Ke, M., Khomutova,
V.I., Kong Z., Kvavadze, E.V., Laarif, F., Lamb, H.E., Lézine, A-M.,
Li, S., Li, W., Liew, P., Liu, G., Liu, J., Liu, Q., Liu, K.-B.,
Lozhkin, A.V., Maley, J., Marchant, R., Mbenza, M., MacDonald, G.M.,
Miyoshi, N., Mock, C.J., Morita, Y., Newby, P., Ni, J., Osipova, I.R.,
Panova, N.K., Perez- Obiol, R., Peyron, O., Prentice, I.C., Qiu, W.,
Reille, M., Ren, G., Reynaud-Farrera, I., Richard, P.J.H., Riollet,
G., Ritchie, J.C., Roche, E., Saarse, L., Scott, L., Sevastyanov,
D.V., Sher, A.V., Song, C., Spear, R.W., Ssemmanda, I., Straka, H.,
Sugita, S., Sun, X., Takahara, H., Tang, L., Tarasov, P.E., Taylor,
D., Thompson, R.S., Uchiyama, T., Van Campo, E., Vilimumbalo, S.,
Vincens, A., Volkova, V.S., Waller, M., Webb III, T., Williams, J.W.,
Xia, Y., Xu, Q., Yan, S., Yang, X., Yu, G., Zernitskaya, V.P., Zhao,
J. and Zheng, Z. The 21 ka Tropical Terrestrial Data Synthesis was
initiated in response to a request by PMIP and compiled by
P.J. Bartlein, R. Bonnefille, M. Bush, I. Farrera, J. Guiot,
S.P. Harrison, K. Holmgren, H. Hooghiemstra, G. Hope, D. Jolly,
S.-E. Lauritzen, Y. Ono, S. Pinot, I.C. Prentice, G. Ramstein,
M. Stute, U. von Grafenstein and G. Yu.
REFERENCES
Bard, E., Rostek, F. and C. Sonzogni, 1997: Interhemispheric synchrony
of the last deglaciation inferred from alkenone
paleothermometry. Nature, 385, 707-710.
Braconnot, P., Harrison, S.P., Hewitt, C.D., Kitoh, A., Otto-Bliesner,
B., Syktus, J., this volume: Preliminary comparison of coupled ocean
atmosphere simulations for 6ka.
Braconnot, P., Joussaume, S., Marti, O and N. de Noblet, 1999:
Synergistic feedbacks from ocean and vegetation on the monsoon
response to mid-Holocene insolation. Geophys. Res. Lett., 26,
2481-2484.
Braconnot, P., Marti, O., Joussaume, S., and Y. Leclainche, in press:
Ocean feedback in response to 6 kyr BP insolation. J. Clim.
Broström, A., Coe, M., Harrison, S.P., Gallimore, R., Kutzbach,
J.E., Foley, J., Prentice, I.C., and P. Behling, 1998: Land surface
feedbacks and paleomonsoons in northern Africa. Geophys. Res. Lett.,
25, 3615- 3618.
Cheddadi, R., Yu, G., Guiot, J., Harrison, S.P. and I.C. Prentice,
1997: The climate of Europe 6000 years ago. Clim. Dyn., 13, 1-9.
Claussen, M., and V. Gayler, 1997: The greening of Sahara during the
mid-Holocene: results of an interactive atmosphere-biome
model. Glob. Ecol. Biogeogr. Lett., 6, 369-377.
Claquin, T., Roelandt, C., Kohfeld, K.E., Harrison, S.P., Prentice,
I.C., Balkanski, Y., Bergametti, G., Hansson, M., Mahowald, N., Rodhe,
N. and M. Schulz, submitted: Radiative forcing of climate by ice-age
dust.
Coe, M.T., 1998: A linked global model of terrestrial hydrologic
processes: simulation of modern rivers, lakes, and
wetlands. J. Geophys. Res., 103, 8885-8899.
Coe, M.T., in press: Modeling terrestrial hydrological systems at the
continental scale: testing the accuracy of an atmospheric
GCM. J. Clim.
Coe, M.T. and S.P. Harrison, this volume: A comparison of the
simulated surface water area in northern Africa for the 6000 yr BP
PMIP experiments.
COHMAP, 1988: Climatic changes of the last 18,000 years: Observations
and model simulations. Science, 241, 1043-1052.
Cowling, S. A., 1999: Simulated effects of low atmospheric
CO2 on vegetation at the Last Glacial Maximum along a North
American latitudinal gradient. Glob. Ecol. Biogeogr. Lett., 8, 81-93.
Cowling, S.A and Sykes, M.T., 1999: Physiological significance of low
atmospheric CO2 for plant-climate interactions. Quat. Res.,
52, 237-242.
de Noblet, N., Claussen, M. and I.C. Prentice, in press: Mid-Holocene
greening of the Sahara: first results of the GAIM 6000 yr BP
experiment with two asynchronously coupled atmosphere/biome
models. Clim. Dyn.
Doherty, R., Kutzbach, J.E., Foley, J.A. and Pollard, D., in press:
Fully-coupled atmosphere/vegetation model simulations of vegetation
feedback effects during the mid- Holocene.
Edwards, M.E., Anderson, P.M., Brubaker, L.B., Ager, T., Andreev,
A.A., Bigelow, N.H., Cwynar, L.C., Eisner, W.R., Harrison, S.P., Hu,
F.-S., Jolly, D., Lozhkin, A.V., MacDonald, G.M., Mock, C.J., Ritchie,
J.C., Sher, A.V., Spear, R.W., Williams, J. and G. Yu, in press:
Pollen-based biomes for Beringia 18,000, 6000 and 0 14C yr
B. P. J. Biogeogr.
Elenga, H., Peyron, O., Bonnefille, R., Prentice, I.C., Jolly, D.,
Cheddadi, R., Guiot, J., Andrieu, V., Bottema, S., Buchet, G., de
Beaulieu, J.L., Hamilton, A.C., Maley, J., Marchant, R., Perez-Obiol,
R., Reille, M., Riollet, G., Scott, L., Straka, H., Taylor, D., van
Campo, E., Vincens, A., Laarif, F. and H. Jonson, in press:
Pollen-based biome reconstruction for southern Europe and Africa
18,000 years ago. J. Biogeogr.
Farrera, I., Harrison, S.P., Prentice, I.C., Ramstein, G., Guiot, J.,
Bartlein, P.J., Bonnefille, R., Bush, M., Cramer, W., von Grafenstein,
U., Holmgren, K., Hooghiemstra, H., Hope, G., Jolly, D., Lauritzen,
S.-E., Ono, Y., Pinot, S., Stute, M. and G. Yu, 1999: Tropical
climates at the last glacial maximum: a new synthesis of terrestrial
palaeoclimatic data. I. Vegetation, lake-levels and
geochemistry. Clim. Dyn., 15, 823-856.
Ganopolski, A., Kubatzki, C., Claussen, M., Brovkin, V. and
V. Petoukhov, 1998: The influence of vegetation-atmosphere-ocean
interaction on climate during the Mid-Holocene. Science, 280,
1916-1919.
Guiot, J., Boreux, J.J., Braconnot, P., Torre, F. and PMIP
Participating Groups, in press. Data- model comparison using fuzzy
logic in palaeoclimatology. Clim. Dyn.
Guiot, J., Structural characteristics of proxy data and methods for
quantitative climate reconstruction, 1991: In: Evaluation of Climate
Proxy Data in Relation to the European Holocene (B. Frenzel, Ed.),
Gustav Fischer Verlag, Stuttgart, 271-284.
Guiot, J., Harrison, S.P. and I.C. Prentice, 1993: Reconstruction of
Holocene precipitation patterns in Europe using pollen and lake-level
data. Quat. Res., 40, 139-149.
Guiot, J., Jolly, D., Cheddadi, R., Peyron, O., Torre, F., Boreux,
J.J. and J.O. Kaplan, this volume: Interpretation of pollen data using
a plant functional type approach and inverse vegetation modelling.
Guiot, J., Torre, F., Jolly, D., Peyron, O., Boreux, J.J. and
R. Cheddadi, submitted: Inverse vegetation modelling: a tool to
reconstruct palaeoclimates under changed CO2 conditions.
Ecol. Model..
Harrison, S.P., Kohfeld, K.E., Roelandt, C., Claquin, T., in press:
The role of dust in climate changes today, at the last glacial maximum
and in the future. Earth Sci. Rev.
Harrison, S.P. and G. Digerfeldt, 1993: European lakes as
paleohydrological and paleoclimatic indicators. Quat. Sci. Rev., 12,
233-248.
Harrison, S.P., Jolly, D., Laarif, F., Abe-Ouchi, A., Dong, B.,
Herterich, K., Hewitt, C., Joussaume, S., Kutzbach, J.E., Mitchell,
J., de Noblet, N. and P. Valdes, 1998: Intercomparison of simulated
global vegetation distribution in response to 6kyr B.P. orbital
forcing. J. Clim., 11, 2721-2742.
Harrison, S.P., Kutzbach, J.E., Prentice, I.C., Behling, P.J. and
M.T. Sykes, 1995: The response of northern Hemisphere extratropical
climate and vegetation to orbitally induced changes in insolation
during the last interglaciation. Quat. Res., 43, 174-184.
Harrison, S.P., Ramstein, G., Braconnot, P., Dong, B., Herterich, K.,
Hewitt, C., Joussaume, S., Kutzbach, J.E., Mitchell, J., Pinot, S.,
Prentice, I.C. and Valdes, P. and PMIP Participating Groups (1999).
Intercomparison of simulated global vegetation in response to 21 kyr
B.P. orbital and glacial forcing. Journal of Climate
Haxeltine, A. and I.C. Prentice, 1996: BIOME3: an equilibrium
terrestrial biosphere model based on ecophysiological constraints,
resource availability, and competition among plant functional
types. Glob. Biogeochem. Cyc., 10, 693-709.
Heimann, M., 1995: The global atmospheric tracer model
TM2. Klimarechenzent, Hamburg, Germany.
Hewitt, C.D. and Mitchell, J.F.B., 1997: Radiative forcing and
response of a GCM to ice age boundary conditions: cloud feedback and
climate sensitivity. Clim. Dyn., 13, 821-834.
Hewitt, C. D. and J.F.B. Mitchell, 1998: A fully coupled GCM
simulation of the climate of the mid-Holocene. Geophys. Res. Lett.,
25, 361-364.
Intergovernmental Panel on Climate Change, Working Group I, 1997:
Technical Paper II: An Introduction to Simple Climate Models Used in
the IPCC Second Assessment Report. World Meteorological Organization,
Geneva, Switzerland.
Jolly, D. and A. Haxeltine, 1997: Effect of low glacial atmospheric
CO2 on tropical African montane vegetation. Science, 276,
786-788.
Jolly, D., Harrison, S.P., Damnati, D. and R. Bonnefille, 1998a:
Simulated climate and biomes of Africa during the Late Quaternary:
Comparison with pollen and lake status data. Quat. Sci. Rev., 17,
629-657.
Jolly, D., Prentice, I.C., Bonnefille, R., Ballouche, A., Bengo, M.,
Brenac, P., Buchet, G., Burney, D., Cazet, J.-P., Cheddadi, R., Edorh,
T., Elenga, H., Elmoutaki, S., Guiot, J., Laarif, F., Lamb, H.,
Lezine, A.-M., Maley, J., Mbenza, M., Peyron, O., Reille, M.,
Reynoud-Farrera, I., Riollet, G., Ritchie, J.C., Roche, E., Scott, L.,
Ssemmanda, I., Straka, H., Umer, M., Van Campo, E., Vilimumbalo, S.,
Vincens, A. and M. Waller, 1998b: Biome reconstruction from pollen and
plant macrofossil data for Africa and the Arabian peninsula at 0 and
6000 years. J. Biogeogr., 25, 1007-1027.
Joussaume, S. and K.E. Taylor, this volume: The Paleoclimate Modeling
Intercomparison Project.
Joussaume, S., Taylor, K.E., Braconnot, P., Mitchell, J.F.B.,
Kutzbach, J.E., Harrison, S.P., Prentice, I.C., Broccoli, A.J.,
Abe-Ouchi, A., Bartlein, P.J., Bonfils, C., Dong, B., Guiot, J.,
Herterich, K., Hewitt, C.D., Jolly, D., Kim, J.W., Kislov, A., Kitoh,
A., Loutre, M.F., Masson, V., McAvaney, B., McFarlane, N., de Noblet,
N., Peltier, W.R., Peterschmitt, J.Y., Pollard, D., Rind, D., Royer,
J.F., Schlesinger, M.E., Syktus, J., Thompson, S., Valdes, P.,
Vettoretti, G., Webb, R.S. and U. Wyputta, 1999: Monsoon changes for
6000 years ago: results of 18 simulations from the Paleoclimate
Modeling Intercomparison Project (PMIP). Geophys. Res. Lett., 26,
859-862.
Kaplan, J., Brubaker, L., Cramer, W., Edwards, M., Harrison, S.P.,
Prentice, I.C., Anderson, P., Andreev, A., Bartlein, P.J., Bigelow,
N., Christensen, T., Lozhkin, A., Matveyeva, N., McGuire, D., Murray,
D., Ritchie, J., Razzhivin, V., Smith, B. and Walker, S., in prep.:
Climatic sensitivity of Arctic terrestrial ecosystem structure and
function: a circumpolar data and modelling synthesis.
Kohfeld, K.E. and S.P. Harrison, 2000: How well can we simulate past
climates? Evaluating the models using global palaeoenvironmental
datasets. Quat. Sci. Rev., 19, 321-346.
Kutzbach, J.E. and Z. Liu, 1997: Response of the African monsoon to
orbital forcing and ocean feedbacks in the Middle Holocene. Science,
278, 440-443.
Kutzbach, J.E. and F.A. Street-Perrott, 1985: Milankovitch forcing of
fluctuations in the level of tropical lakes from 18 to 0 kyr
BP. Nature, 317, 130-134.
Kutzbach, J.E., Bonan, G., Foley, J. and S.P. Harrison, 1996:
Vegetation and soil feedbacks on the response of the African monsoon
to orbital forcing in the early to middle Holocene. Nature, 384,
623-626.
Kutzbach, J.E., Gallimore, R., Harrison, S.P., Behling, B., Selin,
R. and F. Laarif, 1998: Climate and biome simulations for the past
21,000 years. Quat. Sci. Rev., 17, 473-506.
Levis, S., Foley, J.A. and D. Pollard, in press: Climate-vegetation
feedbacks at the Last Glacial Maximum. J.Geophys. Res. (Atmosphere).
Lorenz, S., Grieger, B., Helbig, P. and K. Herterich, 1996:
Investigating the sensitivity of the Atmospheric General Circulation
Model ECHAM 3 to paleoclimatic boundary conditions. Geol. Rundsch.,
85, 513-524.
Mahowald, N., Kohfeld, K.E., Hansson, M., Balkanski, Y., Harrison,
S.P., Prentice, I.C., Rodhe, H. and M. Schulz, 1999: Dust sources and
deposition during the Last Glacial Maximum and current climate: a
comparison of model results with paleodata from ice cores and marine
sediments. J. Geophys. Res., 104, 15,895-16,436.
Mann, M.E., Park, J. and R.S. Bradley, 1995: Global interdecadel and
century-scale climate oscillations during the past five
centuries. Nature, 378, 266-270.
Marticorena, B., and G. Bergametti, 1996: Two-year simulations of
seasonal and interannual changes of the Saharan dust
emissions. Geophys. Res. Let., 23, 1921-1924.
Masson, V., Cheddadi, R., Braconnot, P., Joussaume, S., Texier, D. and
PMIP Participating Groups, 1999: Mid-Holocene climate in Europe: what
can we infer from PMIP model-data comparisons ? Clim. Dyn., 15,
163-182.
Otto-Bleisner, B.L., 1999: El Nino/La Nina and Sahel precipitation
during the middle Holocene. Geophys. Res. Lett., 26, 87-90.
Peterson, G.M., Webb III, T., Kutzbach, J.E., van der Hammen, T.,
Wijmstra, T. and F.A. Street, 1979: The continental record of
environmental conditions at 18,000 yr B. P.: An initial
evaluation. Quat. Res., 12, 47-82.
Peyron, O., Guiot, J., Cheddadi, R., Tarasov, P., Reille, M., de
Beaulieu, J.L., Bottema, S. and V. Andreu, 1998: Climate
reconstruction in Europe for 18 000 yr B.P. from pollen data. Quat.
Res., 49, 183-196.
Pinot, S., Ramstein, G., Harrison, S.P., Prentice, I.C., Guiot, J.,
Joussaume, S. and M. Stute, 1999: Tropical palaeoclimates at the Last
Glacial Maximum: comparison of Paleoclimate Modeling Intercomparison
(PMIP) simulations and paleodata. Clim. Dyn., 15, 857-874.
Prentice, I.C., Guiot, J. and S.P. Harrison, 1992: Mediterranean
vegetation, lake levels and palaeoclimate at the Last Glacial
Maximum. Nature, 360, 658-670.
Prentice, I. C., and T. Webb III, 1998: BIOME 6000: reconstructing
global mid-Holocene vegetation patterns from palaeoecological
records. J. Biogeogr., 25, 997-1005.
Prentice, I.C., Cramer, W., Harrison, S.P., Leemans, R., Monserud,
R.A. and A.M. Solomon (1992). A global biome model based on plant
physiology and dominance, soil properties, and climate. J. Biogeogr.,
19, 117-134.
Prentice, I.C., Guiot, J., Huntley, B., Jolly, D. and R. Cheddadi,
1996: Reconstructing biomes from palaeoecological data: a general
method and its application to European pollen data at 0 and 6
ka. Clim. Dyn., 12, 185-194.
Prentice, I.C., Jolly, D. and BIOME 6000 Members, in press:
Mid-Holocene and Glacial Maximum vegetation geography of the northern
continents and Africa. J. Biogeogr.
Qin, B., Harrison, S.P. and J.E. Kutzbach, 1998: Evaluation of
modelled regional water balance using lake status data: a comparison
of 6ka simulations with the NCAR CCM. Quat. Sci. Rev., 17, 535-548.
Rind, D. and D. Peteet, 1985: Terrestrial conditions at the last
glacial maximum and CLIMAP sea-surface temperature estimates: are they
consistent? Quat. Res., 24, 1-22.
Rosell-Melé, A., Bard, E., Emeis, K.C., Farrimond, P., Grimalt,
J., Muller, P.J. and Schneider, R.R., 1998: Project takes a new look
at past sea surface temperatures. EOS Trans. of AGU, 79, 393-394.
Rostek, F., Ruhland, G., Bassinot, F.C., Muller, P.J., Labeyrie, L.D.,
Lancelot, Y. and E. Bard, 1993: Reconstructing sea-surface temperature
and salinity using ?18O and alkenone records. Nature, 364, 319-321.
Schneider, R.R., Mueller, P.J. and G. Ruhland, 1995: Late Quaternary
surface circulation in the east-equatorial South Atlantic: Evidence
from alkenone sea surface temperatures. Paleoceanography, 10,
197-219.
Schulz, M., Balkanski, Y., Guelle, W. and F. Dulac, F. 1998: Role of
aerosol size distribution and source location in a three-dimension
simulation of a Saharan dust episode tested against satellite-derived
optical thickness. J. Geophys. Res., 103, 10,579-10,592.
Sonzogni, C., Bard, E. and F. Rostek, 1998: Tropical sea-surface
temperatures during the last glacial period: a view based on alkenones
in Indian Ocean sediments. Quat. Sci. Rev., 17, 1185-1201.
Steffen, W.L., Walker, B.H., Ingram, J.S. and G.W. Koch, 1992: Global
change and terrestrial ecosystems: The operational plan. Global Change
Report 21.
Street, F.A. and A.T. Grove, 1976: Environmental and climatic
implications of late Quaternary lake-level fluctuations in
Africa. Nature, 261, 385-390.
Street-Perrott, F.A. and S.P. Harrison, 1985: Lake levels and climate
reconstruction. In Paleoclimate Analysis and Modeling. (A. D. Hecht,
Ed.), pp. 291-340. John Wiley, New York.
Street-Perrott, F.A., Huang, Y., Perrott, R.A., Eglington, G., Barker,
P., Khelifa, L.B., Harkness, D.D., and D.O. Olago, 1997: Impact of
lower atmospheric carbon dioxide on tropical mountain
ecosystems. Science, 278, 1422-1426.
Street-Perrott, F.A., Marchand, D.S., Roberts, N. and S.P. Harrison,
1989: Global lake-level variations from 18,000 to 0 years ago: a
palaeoclimatic analysis. U. S. Department of Energy, Washington, DC.
Tarasov, P.E., Guiot, J., Cheddadi, R., Andreev, A.A., Bezusko, L.G.,
Blyakharchuk, T.A., Dorofeyuk, N.I., Filimonova, L.M., Volkova,
V.S. and V.P. Zernikskaya, in press: Climate in northern Eurasia 6000
years ago reconstructed from pollen data. Earth and Planetary Science
Letters.
Tarasov, P.E., Harrison, S.P., Saarse, L., Pushenko, M.Ya., Andreev,
A.A., Aleshinskaya, Z.V., Davydova, N.N., Dorofeyuk, N.I., Efremov,
Yu.V., Khomutova, V.I., Sevastyanov, D.V., Tamosaitis, J., Uspenskaya,
O.N., Yakushko, O.F. and I.V. Tarasova, 1994: Lake status records from
the former Soviet Union and Mongolia: Data base documentation. NOAA
Paleoclimatology Publications Series Report, 2, 274pp.
Tarasov, P.E., Pushenko, M.Y., Harrison, S.P., Saarse, L., Andreev,
A.A., Aleshinskaya, Z.V., Davydova, N.N., Dorofeyuk, N.I., Efremov,
Y.V., Elina, G.A., Elovicheva, Y., Filimonova, L.V., Gunova, V.S.,
Khomutova, V.I., Kvavadze, E.V., Neustreuva, I., Pisareva, V.V.,
Sevastyanov, D.V., Shelekhova, T.S., Subetto, D.A., Uspenskaya,
O.N. and V.P. Zernitskaya, 1996: Lake status records from the former
Soviet Union and Monglia: Documentation of the second version of the
database. NOAA Paleoclimatology Pulbications Series Report, 5, 224pp.
Tarasov, P.E., Peyron, O., Guiot, J., Brewer, S., Volkova, V.S.,
Bezusko, L.G., Dorofeyuk, N.I., Kvavadze, E.V., Osipova, I.M. and
N.K. Panova, 1999: Last Glacial Maximum climate of the former Soviet
Union and Mongolia reconstructed from pollen and plant macrofossil
data. Clim. Dyn., 15, 227-240.
Tarasov, P.E., Webb III, T., Andreev, A.A., Afanas'eva, N.B.,
Berezina, N.A., Bezusko, L.G., Blyakhararchuk, T.A., Bolikhovskaya,
N.S., Cheddadi, R., Chernavskaya, M.M., Chernova, G.M., Dorofeyuk,
N.I., Dirksen, V.G., Elina, G.A., Filimonova, L.V., Glebov, F.Z.,
Guiot, J., Gunova, V.S., Harrison, S.P., Jolly, D., Khomutova, V.I.,
Kvavadze, E.V., Osipova, I.M., Panova, N.K., Prentice, I.C., Saarse,
L., Sevastyanov, D.V., Volkova, V.S. and V.K. Zernitskaya, 1998:
Present-day and mid-Holocene biomes reconstructed from pollen and
plant macrofossil data from the former Soviet Union and
Mongolia. J. Biogeogr., 25, 1029-1053.
TEMPO, 1996: Potential role of vegetation feedbacks in the climate
sensitivity of high-latitude regions: A case study at 6000 years
B.P. Glob. Biogeochem. Cyc., 10, 727-736.
Tett, S.F.B., Stott, P.A., Allen, M.R., Ingram, W.J. and
J.F.B. Mitchell, 1999: Causes of twentieth-century temperature change
near the Earth's surface. Nature 399, 569-572.
Texier, D., de Noblet, N. and P. Braconnot, 2000: Sensitivity of the
African and Asian monsoons to mid-Holocene insolation and
data-inferred surface changes. J. Clim., 13, 164-181.
Texier, D., de Noblet, N., Harrison, S.P., Haxeltine, A., Jolly, D.,
Joussaume, S., Laarif, F., Prentice, I.C. and P. Tarasov, 1997:
Quantifying the role of biosphere-atmosphere feedbacks in climate
change: Coupled model simulation for 6000 years BP and comparison with
palaeodata for northern Eurasia and northern Africa. Clim. Dyn., 13,
865-882.
Thompson, R.S. and K.H. Anderson, in press: Biomes of Western North
America at 18,000, 6,000, and 0 14C yr B.P. reconstructed
from pollen and packrat midden data. J. Biogeogr.
Vettoretti, G., and Peltier, W.R. (1998). Simulations of Mid-Holocene
climate using an atmospheric general circulation model. J. Clim., 11,
2607-2627.
Webb III, T., Bartlein, P.J., Harrison, S.P. and K.H. Anderson, 1993:
Vegetation, lake levels and climate in eastern North America for the
past 18,000 years. In Global Climates since the Last Glacial
Maximum. (H.E. Wright Jr., J.E. Kutzbach, T. Webb III, W.F. Ruddiman,
F.A. Street-Perrott, and P.J. Bartlein, Eds.),
pp. 415-467. University of Minnesota, Minneapolis, MN.
Williams, J.W., Webb III, T., Richard, P.J.H. and P. Newby, in press:
Late Quaternary biomes of Canada and the eastern United
States. J. Biogeogr.
Wright Jr., H.E., Kutzbach, J.E., Webb III, T., Ruddiman, W.F.,
Street-Perrott, F.A. and P.J. Bartlein, 1993: Global Climates since
the Last Glacial Maximum. University of Minnesota Press, Minneapolis,
MN.
Yu, G. and S.P. Harrison, 1996: An evaluation of the simulated water
balance of Eurasia and northern Africa at 6000 yr BP using lake status
data. Clim. Dyn., 12, 723-735.
Yu, G., Bin, X. and J. Liu, this volume: Synthesis of
palaeoenvironmental evidence at 6ka and 21ka and reconstructions of
the Asian palaeo-monsoon changes.
Yu, G., Chen, X., Ni, J., Cheddadi, R., Guiot, J., Han, H., Harrison,
S.P., Huang, C., Ke, M., Kong, Z., Li, S., Li, W., Liew, P., Liu, G.,
Liu, J., Liu, Q., Liu, K.-B., Prentice, I.C., Qui, W., Ren, G., Song,
C., Sugita, S., Sun, X., Tang, L., Campo, E.V., Xia, Y., Xu, Q., Yan,
S., Yang, X., Zhao, J. and Z. Zheng, in press: Palaeovegetation of
China: a pollen data-based synthesis for the mid-Holocene, the last
glacial maximum. J. Biogeogr.
Yu, G., Prentice, I.C., Harrison, S.P. and X. Sun, 1998: Pollen-based
biome reconstructions for China at 0 ka and 6 ka. J. Biogeogr., 25,
1055-1069.
Yu, G. and B. Qin, 1997: Holocene temperature and precipitation
reconstructions and monsoonal climates in eastern China using pollen
data. Paleoclimates, 2, 1-32.
|
| Home |
Top |
Last updated 2001/10/09 15:09:03 |
 (Click on the figure to
get a bigger version)
(Click on the figure to
get a bigger version)